Om planter, dyr, energi og de eldste organismer
 Uten plantene ville det ikke ha vært noe dyreliv på jorda. Ved basis av dyrenes næringskjede står de som spiser planter. Alle levende organismer må ha tilførsel av energi for å holde livsprosessen i gang. Dyrene kan bare få energi ved å spise energirike matemner som kommer fra andre organismer. Plantene, derimot, fanger inn lysenergi i de grønne delene (klorofyll) og binder den til energirike organiske forbindelser som de lager (syntetiserer) fra de enkle forbindelsene vann (H2O) og karbondioksid (CO2). Prosessen kalles fotosyntese (photos, gresk = lys). Vi sier at plantene er autotrofe og dyrene heterotrofe (se tekstboks). Plantene overfører lysenergi til kjemisk bundet energi som heterotrofe organismer igjen kan frigjøre og benytte i livsprosessen.
Uten plantene ville det ikke ha vært noe dyreliv på jorda. Ved basis av dyrenes næringskjede står de som spiser planter. Alle levende organismer må ha tilførsel av energi for å holde livsprosessen i gang. Dyrene kan bare få energi ved å spise energirike matemner som kommer fra andre organismer. Plantene, derimot, fanger inn lysenergi i de grønne delene (klorofyll) og binder den til energirike organiske forbindelser som de lager (syntetiserer) fra de enkle forbindelsene vann (H2O) og karbondioksid (CO2). Prosessen kalles fotosyntese (photos, gresk = lys). Vi sier at plantene er autotrofe og dyrene heterotrofe (se tekstboks). Plantene overfører lysenergi til kjemisk bundet energi som heterotrofe organismer igjen kan frigjøre og benytte i livsprosessen.
Når plantene spiller en slik sentral rolle i overføringen av energi, forstår vi at plantelivets utvikling må ha hatt avgjørende betydning for utviklingen av dyrelivet på jorda. Det gjelder særlig plantenes utvikling siden de begynte å kolonisere tørt land for ca. 450 millioner år siden. Det er den delen av plantenes utvikling vi vet mest om gjennom fossiler.
I jordas tidlige historie var den ultrafiolette (UV-) strålingen sterk fordi atmosfæren manglet ozonlaget som reduserer denne strålingen. Fordi UV-strålingen dreper levende celler, er det logisk å anta at de tidligste éncellete organismene utviklet seg på dypt vann hvor de var skjermet mot strålingen. På dypt vann er det ikke lys og mulighet for fotosyntese. Det antas derfor at de tidligste autotrofe organismene må ha hatt kjemosyntese, som skiller seg fra fotosyntesen ved at den bruker kjemisk bundet energi i stedet for lysenergi (mer om dette i tekstboksen «Autotrofe og heterotrofe organismer»).
I jordas opprinnelige atmosfære var det lite oksygen. Under fotosyntesen blir vannmolekyler spaltet og oksygen (O2) frigjort. Fotosyntesen økte derfor etterhvert oksygeninnholdet i atmosfæren og det ble dannet et ozonlag (O3) som reduserer UV-strålingen fra sola. I jordas tidlige historie var temperaturen i atmosfæren temmelig høy på grunn av store mengder drivhusgasser, bl.a. CO2. Med fotosyntesen ble også dette endret, da mye CO2 ble bundet i organiske forbindelser i levende organismer og i fossilt organisk materiale (kull, olje, gass og finfordelt i sedimenter).
De tidligste éncellete organismene og senere de flercellete algene besto av bløtt og lett forgjengelig materiale. Det finnes derfor sparsomt med fossiler etter dem. De eldste fossiler som er kjent, er funnet i ca. 3,6 milliarder (mrd) år gamle bergarter i Australia og på Grønland. Disse er beslektet med de blågrønne bakteriene (cyanobakterier) som mangler cellekjerne (prokaryoter, se systematisk oversikt bakerst). Fossile bakterier er kjent fra 2 mrd år gamle bergarter. Éncellete organismer med ekte cellekjerne (eukaryoter) er kjent fra ca. 1,8 (muligens 2,1) mrd år gamle avsetninger i Nord-Amerika. I dolomittbergarter i Finnmark finnes stromatolitter (se tekstboks) av tilsvarende alder. I prekambriske lag (se geologisk tidsskala) er akritarker (se tekstboks) ganske vanlige. Fossiler av flercellete alger er funnet i bergarter som er omkring 800 millioner år gamle, bl.a. fra Svalbard.
|
Autotrofe og heterotrofe organismer
Autotrof og heterotrof er betegnelser på ulike måter som organismene skaffer seg energi på til livsprosessene (auto, gresk = selv; trof, fra gresk trophe = mat; hetero, gresk = forskjellig). En autotrof organisme er "selvmatende", det vil si den kan danne energirike, organiske forbindelser fra enkle, uorganiske forbindelser (karbondioksid og vann) ved bruk av energi fra omgivelsene. I fotosyntesen er energien sollys, i kjemosyntesen er den bundet i uorganiske molekyler og frigjøres ved oksidasjon, men ellers er kjemosyntesen helt lik fotosyntesen. I begge syntesetypene brukes energien til å spalte vannmolekyler (H2O) og bygge hydrogenet sammen med karbondioksid (CO2) til energirike karbohydratmolekyler. Alle grønne planter og andre eukaryoter som har klorofyll (planktonalger) er autotrofe og har fotosyntese. De aller fleste bakterier og alle dyr er heterotrofe; de kan ikke produsere sin egen næring ved hjelp av energi fra omgivelsene, men må fortære andre organismer for å skaffe seg energi. Noen få bakterieformer har imidlertid kjemosyntese og er autotrofe; eksempler er svovelbakterier som får energi til syntesen ved å oksiderer hydrogensulfid til svovel og videre til sulfat (H2S -> S ® SO4- - ) og jernbakterier som oksiderer toverdig jern til treverdig jern (Fe2+ -> Fe3+). I 1977 oppdaget en bemannet ubåt svovelrike varmtvannsutstrøminger på ca. 2500 m dyp utenfor Guatemala, hvor det lever autotrofe bakterier som danner livsgrunnlaget for et mangfoldig dyreliv.De eldste livsformene på jorda var sannsynligvis lignende former av autotrofe bakterier, og de fleste biologer tror at fotosyntesen ble utviklet hos organismer med kjemosyntese som utviklet klorofyll.
|
|
Akritarker
Akritarker hører til de eldste kjente mikrofossiler og finnes i marine avsetninger fra tidlig i jordas urtid og fram til i dag. Den biologiske opprinnelsen er usikker, men en regner at de er hvilesporer eller cyster, for en del sannsynligvis fra planktoniske algeformer. De var kuleaktige eller mer eller mindre kantete hylstre med pigger og utvekster av ulike slag. Da de finnes i paleozoiske og prekambriske bergarter hvor det er lite andre fossiler, kan de være til hjelp ved aldersbestemmelse.
|
Makrofossiler og mikrofossiler
Makrofossiler kan sees uten å bruke lupe eller mikroskop, men for nærmere undersøkelser brukes mikroskop. Mikrofossiler er fossiler som bare kan sees og studeres under sterk forstørrelse, og det må som regel benyttes spesielle prepareringsteknikker for å frigjøre dem fra bergarten de er oppbevart i. Plantemakrofossiler er svært sjelden hele planter, men avtrykk eller kullaktige rester av blad og stengler som oftest er mer eller mindre fragmentert. Fossile frø, frukter og andre fruktifikasjoner regnes som makrofossiler. Sporer og pollen (blomsterstøv) er plantemikrofossiler; akritarker og éncellete alger opptrer også som mikrofossiler. Det kan være titusener til hundretusener vel bevarte mikrofossiler i ett gram sediment, mens store antall av gode makrofossiler ikke er så vanlig. Fossilførende bergarter inneholder vanligvis både makro- og mikrofossiler.
|
Stromatolitter
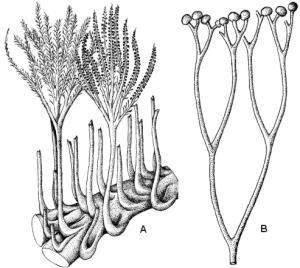
Tueliknende til søyleformete kalkdannelser med lagdeling som er kjent fra tidlig prekambrium. Stromatolitter dannes i dag av matter av trådformete blågrønne bakterier (cyanobakterier) og grønnalger i tidevannssonen i grunne havområder. De slimete algetrådene fanger opp kalkslam som kittes sammen til lagdelte kalkstrukturer. Lagdelingen skyldes variasjoner i algenes tilvekst. Stromatolitter fra Raipasgruppen i Alta kan være 1950 - 2100 millioner år gamle og hører til de eldste spor etter liv vi kjenner i Norge.
|
De første landplantene
Det neste store steg i livsformenes utviklingshistorie etter at livet var oppstått på jorda, var utviklingen av planter som kunne overleve på tørt land. Ettersom planter danner næringsgrunnlaget for dyr, er det rimelig å anta at plantene inntok landjorda før dyrene. Før plantenes kolonisering av tørt land hadde éncellete og flercellete algeformer inntatt havområdene i omtrent samme omfang som i dag.
Utviklingen fra et liv i vann til et liv på land omgitt av luft førte til mange forandringer og tilpasninger hos plantene. De utviklet et beskyttende hudlag med et vokslignende belegg (kutikula) som nedsetter fordampningen. Men dette laget hindrer også passasjen av karbondioksid som er nødvendig for fotosyntesen. Som en løsning på dette utviklet de «ventiler» i hudvevet (spalteåpninger) som kan stenges når vanntapet blir stort. Videre utviklet de organer som tar opp vann og næringssalter fra jorda (røtter), og ledningsvev som transporterer det til plantedelene over jorda. Ledningsvevet virker også som støtte for plantekroppen så den kan reise seg opp fra marka og danne grener for å få mest mulig lys.
En av de viktigste tilpasningene plantene måtte gjennom for å kunne leve på land, gjelder den kjønnete formeringen. Organismer som lever i vann, kan slippe kjønnscellene direkte ut i vannet og la befruktningen foregå der. Også på land må kjønnscellene være omgitt av vann for å kunne fylle sin funksjon. De kan ikke overleve uttørking.
Fra sine forgjengere blant algene arvet landplantene en livssyklus med to atskilte generasjoner, én med dobbelt antall kromosomer (diploid), og én med enkelt antall kromosomer (haploid) hvor kjønnscellene blir dannet. Sammensmeltningen av en hannlig og en hunnlig kjønnscelle (sædcelle og eggcelle) starter en ny, ukjønnet plante som er diploid. Vann, som er nødvendig for at sædceller og eggceller skal produseres og møtes, finnes på land som regn, dogg og andre former for tilfeldig fuktighet i omgivelsene. For å sikre at et tilstrekkelig antall av den kjønnete generasjon skal treffe slike omgivelser, har landplantene sørget for vid spredning av et stort antall sporer. Sporene er karsporeplantenes kjønnete generasjonsenheter. De er mindre enn 0,1 mm og produseres i enorme antall av hvert planteindivid. Som beskyttelse mot uttørking har de en vanntett ytterhinne (exine). Sporene føres lett av sted med vinden, slik at det er stor sjanse for at noen vil lande i gunstige omgivelser. I plantenes videre utvikling skjedde de største endringene nettopp i måten den kjønnete generasjonen ble tatt vare på. Hos frøplantene blir eggcellene beholdt i blomsten, som også sørger for et miljø der de hannlige kjønnscellene (pollen) kan befrukte eggcellen uavhengig av tilfeldig vann fra omgivelsene.
Lenge før utviklingen av landplanter hadde en del alger tilpasset seg et liv på land i fuktige miljøer med periodevis uttørking, men de kunne ikke danne noe grønt plantedekke utenom de spesielle miljøene. Alger mangler celler som kan ta opp vann fra jorda og ledningsceller som kan transportere vann og næring i plantekroppen. Før landplantene kom, var derfor jordoverflaten naken og gold som en ørken, unntatt på fuktige steder hvor det kunne lyse grønt i en tynn algevegetasjon. Det er mulig at det fantes lav. De består av nokså primitive organismer, sopp og éncellete alger, som lever sammen i symbiose og danner en slags planter. Men vi kjenner ingen fossiler som kan si oss noe om lavenes alder.
Med utviklingen av et ledningsvev fikk vi det som kan kalles de første ekte landplantene, - karplantene (kar = ledningsceller). Vi vet ikke sikkert når de første oppsto. De eldste kjente makrofossilene (se tekstboks) er fra siste del av silur. Sporer, som ser ut til å være dannet av høyere planter, er funnet så tidlig som midtre ordovicium. De gjør det sannsynlig at ekte landplanter begynte å utvikles en gang for omkring 470 millioner år siden.
Moser
Mosene står i en mellomstilling mellom alger og karplanter når det gjelder deres forhold til vann. De har ikke røtter eller tilsvarende organer for vannopptak fra jorda, og de har ikke ledningsvev som karplantene. De er derfor avhengig av et fuktig miljø på samme måte som algeformer som finnes på land, og overlever uttørking ved «å gå i dvale». Likevel kan de danne et tett og sammenhengende plantedekke, men de mangler vev som kan gi dem stivhet til å vokse i høyden. Det har vært spekulert på om karplantene kan ha utviklet seg fra moser. De eldste makrofossiler av moser er fra silur, nesten like gamle som de eldste karplanter vi kjenner.
Den egentlige moseplanten er den haploide, kjønnete generasjonen, mens hos karplantene er den kjønnete generasjonen svært kortvarig og beskjeden i forhold til den egentlige planten, som er diploid. Dette har gitt karplantene et fortrinn i utviklingen av ulike tilpasninger til livet på land. Mosene ansees som et «sidespor» i utviklingen, hvor hovedvekten er lagt på den haploide generasjonen som mangler potensiale for å utvikles videre til høyere planter.
|

|
|
|
Nålevende bjørnemose
|
A: Hyenia (10-20 cm høy); B: Cooksonia (6 cm høy)
|
 |
Karsporeplanter
De eldste kjente karplantene produserte sporer til spredning av kjønnsgenerasjonen og kalles derfor karsporeplanter. Karsporeplantene omfatter i dag bregner (ca. 10 000 arter), kråkefotplanter (ca. 1000 arter) og snelleplanter (25 arter) og er den eldste gruppen av nålevende høyere planter. Makrofossiler av sikre karplanter er ikke funnet i lag eldre enn silur. Den eldste er Cooksonia, som ble ca. 6 cm høy. Ved begynnelsen av devonperioden, for ca 400 millioner år siden, hadde det utviklet seg et stort antall urteformete karsporeplanter, bl.a. Zosterophyllum som kråkefotplantene har utviklet seg fra, og Rhynia som er utgangspunktet for bregnene og snelleplantene (kjerringrokk). Disse plantene hadde ikke ekte røtter, men krypende jordstengler med utvekster som kunne ta opp vann; egentlige blad hadde de heller ikke, men noen hadde skjellformete utvekster som kan ha fungert som blad.
De tre gruppene av karsporeplanter som lever i dag, fantes allerede i devon, og mot slutten av devonperioden hadde de utviklet store trær. De fortsatte gjennom karbon hvor de var dominerende i sumpskogene som ble omdannet til kull, særlig skjelltrærne Lepidodendron og Sigillaria og snelletrærne Calamites. Noen av dem var 30-50 meter høye. Psaronius er eksempel på en stor trebregne. De treformete kråkefot- og snelleplantene døde ut i perm; de nålevende artene er alle urter. Blant bregnene er det fortsatt noen som er trær.
Til venstre: Skjelltreet Sigillaria som kunne bli 30 m høyt.
|
Frøplanter: Nakenfrøete (gymnospermer) og dekkfrøete (angiospermer)
Frøplantene skiller seg fra sporeplantene ved at de hunnlige kjønnscellene (eggcellene) ikke slipper fri fra morplanten, men er plassert i frøemner som er samlet i kongler, blomster eller tilsvarende organer. De hannlige kjønnscellene blir spredt med pollenet, som svarer til hannlige sporer hos karsporeplantene. Hos gymnospermene er frøemnene ubeskyttet (gymnos, gresk = naken), slik at pollenet kan lande direkte på frøemnet. Hos angiospermene er frøemnet omgitt av en beholder (fruktknuten; angeios, gresk = beholder) som modnes til frukt; gymnospermene har ikke frukt, bare frø.
Utviklingen av frøet og deretter av fruktknuten var de to viktigste trinn i plantenes utvikling etterat de hadde kolonisert landjorda. Frøet er en liten kimplante som er utstyrt med frønæring, en «matpakke» den forbruker under spiringen inntil den har slått rot og kan klare seg selv. Dette var et viktig skritt for sikring av avkommet. Utviklingen av frø og senere frukter resulterte i et vell av spesialiserte tilpasninger for effektiv spredning med vind og med dyr.
De første frøplantene, som var gymnospermer, oppsto i den midtre delen av devonperioden, for ca. 370-380 millioner år siden. Angiospermene oppsto i begynnelsen av krittperioden, for ca. 140 millioner år siden, og hadde en rask utvikling og spredning.
Gymnospermenes utvikling i paleozoikum
De tidlige gymnospermene hadde bregnelignende blad og kalles derfor frøbregner. Noen var trær, som for eksempel Medullosa. Men de fleste var lianer, det vil si klatreplanter, særlig i karbonperiodens sumpskoger av karsporeplanter. Cordaiter var en annen og viktig gruppe i karbon. De kunne bli opptil 50 m høye trær som hadde flere desimeter lange, tykke blad og dannet skoger. Mot slutten av karbonperioden kom også bartrær og cycadéer. Alle disse gymnospermene fortsatte inn i permperioden, hvor bartrærne etter hvert spilte en stadig større rolle. Karbonperioden har fått sitt navn på grunn av at de største kullfeltene i verden ble dannet av enorme sumpskoger i denne perioden.
|
Kull
Naturlig kull er en karbonrik, brennbar bergart som er dannet av planterester; trekull er ingen bergart, men laget ved forkulling av ved. Utgangsmaterialet for naturlige kull er torv eller lignende konsentrasjoner av planterester. For å bli omdannet til kull må planterestene bli overleiret av flere hundre til noen tusen meter tykke sedimenter og bli utsatt for jordvarme. I de øvre ti tusen meter av jordskorpen øker temperaturen gjennomsnittlig ca. 3º C for hver hundre meter. De opprinnelige planterestene består av ca. 60 % karbon, resten er mest oksygen og hydrogen, og mindre mengder for eksempel nitrogen og svovel. Når det lages trekull, skjer brenningen under høy temperatur og uten oksygentilførsel, slik at alt unntatt karbon forbrenner fullstendig; trekull er praktisk talt rent karbon. Dannelsen av naturlige kull tar millioner av år under temperaturer fra 50º til litt over 100º C. Hvilken type av kull som blir resultatet, brunkull, steinkull eller antrasitt, avhenger av hvor høy temperaturen har vært. Noen få hundre meter overleiring gir svak temperaturstigning og dannelse av brunkull; overleiring på 1500-3000 m og temperatur 75-100º fører til steinkull; større overleiring fører til høyere temperatur og dannelse av antrasitt. Karboninnholdet i naturlige kulltyper øker fra 65% til 98% fra brunkull til antrasitt; torv har omkring 60% karbon. I motsetning til trekull inneholder naturlige kull også oksygen og hydrogen, som er bundet til karbon i komplekse molekyler som ingen enda har vært i stand til å analysere i detalj. Et av de viktigste plantestoffene i kulldannelsen er lignin. Ved består av ca. 2/3 cellulose og resten lignin. Cellulose brytes lett ned av mikroorganismer, mens lignin blir ufullstendig nedbrutt og virker hemmende på nedbrytningen av andre plantestoffer. Det er en viktig bestanddel i det vi kaller humus, som har en komplisert og ufullstendig kjent kjemi. Under dannelsen av kull blir lignin og humus omdannet til stadig større og mer kompliserte molekyler som utgjør hoveddelen av kullmassen.
På Spitsbergen tar norske og russiske gruver ut tertiære kull, dannet for 50-60 millioner år siden, mens en tidligere russisk gruve i Pyramiden var basert på kull dannet i karbontiden, for ca 300 millioner år siden. På Bjørnøya ble det tidligere tatt ut devonske kull.
|
Gymnospermenes utvikling etter paleozoikum
I størstedelen av mesozoikum var jordas vegetasjon dominert av gymnospermer. Det var bartrær, konglepalmer (cycadéer), bennettitter, ginkgoer og frøbregner som dannet trevegetasjonen, med bregner og andre karsporeplanter som viktige innslag i skogenes undervegetasjon og i åpne landskap. I krittperioden fikk disse plantene en sterk konkurrent i angiospermene, som fra midten av perioden ble den dominerende plantegruppen overalt på jorda. Bennettitter og frøbregner forsvant helt, ginkgoer ble redusert til noen ganske få, slik at i dag er det bare én art igjen av denne gruppen: Ginkgo biloba, det kinesiske tempeltreet. Cycadéene gikk sterkt tilbake; de kalles konglepalmer fordi de har blad som minner om palmer. I dag finnes det ca. 100 arter i tropiske og subtropiske strøk. Bartrærne var den gruppen av gymnospermer som klarte seg best i konkurransen med angiospermene. I dag finnes det ca. 600 arter av bartrær, men deres rolle i jordens vegetasjon er større enn dette antallet skulle tilsi. Mange arter danner sammenhengende skoger over store områder i middelsvarme strøk på den nordlige og sydlige halvkule. I mesozoikum var det langt flere arter av bartrær, men de spilte neppe noen større rolle i jordens samlete vegetasjon enn de gjør i dag.
 |
 |
|
Fossile Ginkgo-blader
|
Nålevende Ginkgo biloba fra Botanisk hage, Oslo
|
 |
Angiospermer - dekkfrøete blomsterplanter
Framveksten av angiospermene i løpet av krittperioden (145-65 millioner år siden) var en revolusjon i utviklingen av plantelivet på jorda. De utkonkurrerte storparten av de eldre plantegruppene og har de siste 60-70 millioner år vært helt dominerende. I dag er det i overkant av 250 000 arter av angiospermer, dvs. ca. 95% av alle artene av høyere planter som lever på jorda.
Angiospermene utviklet seg sannsynligvis fra en spesialisert gruppe av frøbregner. Det kan ikke pekes på globale klimatiske eller andre miljømessige omveltninger som årsak til deres raske framvekst. Forklaringen ligger heller i utviklingen av blomsterorganene og deres spesielle form for befruktning som ga dem et overlegent potensiale for utvikling av variasjon i spredningsmåter for pollen og for frø og frukter.
|
|
Magnolia (fra Botanisk hage, Oslo) tilhører en familie
av dekkfrøete planter som er kjent som fossil helt tilbake til kritt.
|
|
 |
Falske plantefossiler: dendritter
På sprekkflater i stein finner en ofte moselignende forgrenete figurer som kan forveksles med plantefossiler. De er dannet ved at bl.a. jern og mangan har blitt felt ut fra vann som har passert gjennom sprekkene; metallsaltene har krystallisert på sprekkflatene i steinen i likhet med isroser på en vindusflate.
|
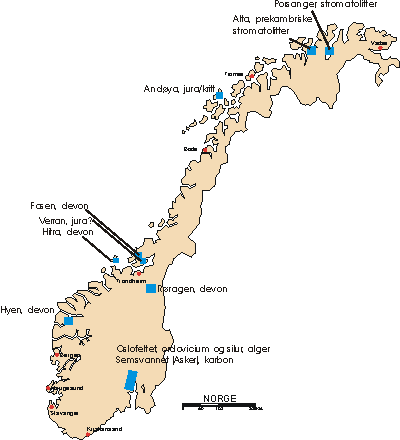
Plantefossiler i Norge (lokaliteter er vist på kartet)
 |
|
Kart som viser steder i Norge hvor det er funnet plantefossiler.
|
 |
Alger
Alger etterlater sjelden fossiler fordi de er bløte og lett forgjengelige; unntaket er kalkalger som skiller ut kalk i og på celleveggene så de på en måte blir «forsteinet» mens de lever. Kalkalger er ikke noen egen systematisk gruppe, men forskjellige grønnalger og rødalger som skiller ut kalk (forkalkete brunalger er ukjent). De utgjør bare en liten del av de totale antallet algearter.
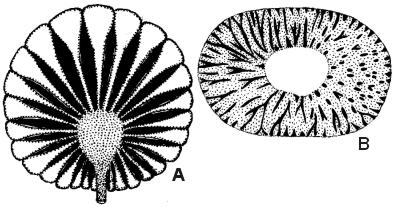
Oslofeltets kambro-silur-bergarter inneholder mange kalkalger. I ordovicium-lagene er de kuleformete typene Coelosphaeridium (A på tegningen, tverrsnitt = 3-4 cm) og Cyclocrinus svært vanlige. I silurlagene opptrer bl.a. Mastopora, som var mindre forkalket og derfor presset til en mørk sirkelformet flate med et sekskantet cellemønster. Den sigarformete Paleoporella (B på tegningen, tverrsnitt = 3-5 mm) er vanlig i de øverste ordoviciumlagene; ved Tyrifjorden er det 20 meter tykke kalklag der denne arten er enerådende.
|
Høyere planter
I Norge er det lite med avsetninger fra den delen av jordas historie da det har eksistert landplanter. Det meste er fra devonperioden, og ellers er det noen små forekomster fra karbon og jura.
Devon
Landplanter fra devonperioden finnes i Røragenområdet mellom Røros og grensen til Sverige. En lignende devonflora finnes langs Trøndelagskysten, og en litt yngre finnes på sørsiden av Nordfjord på nord-vestlandet. Floraene består av typiske under- til mellomdevonplanter med Psilophyton som en ganske vanlig form. Hyenia er en planteslekt som har fått navn etter Hyen i Nordfjord hvor den først ble funnet og beskrevet. Fossilene er relativt dårlig oppbevart.
Karbon
Ved Semsvann i Asker finnes det plantefossiler fra siste del av karbonperioden i sandsteiner og skifre. De vanligste fossilene er Calamites, som er slektninger av dagens snelleplanter. Det er videre funnet blad av frøbregner, blad og frø av Cordaites og skudd av bartrær.
Jura
I 1867 ble det oppdaget kullag på Andøya nær Skarstein, men de var ikke økonomisk drivverdige. Skiferlagene inneholder til dels ganske mye plantefossiler, særlig bartrenåler. Lagene inneholder dessuten mye pollen og sporer. Makro- og mikrofossilene viser en typisk flora fra juratiden av bregner, bartrær, konglepalmer, bennettitter og ginkgoer.
I Verran ved Beitstadfjorden i Nord-Trøndelag ble det i 1920-årene funnet kull som etter alt å dømme var fra stedet, men det var ikke mulig å finne dem i fast fjell. Kullene inneholder sporer, pollen og bladfragmenter (kutikula) som viser at de er fra juraperioden.
Geofysiske undersøkelser har vist at lag fra juraperioden finnes i bergrunnen i fjorden utenfor.
Plantefossiler på Svalbard
Svalbard har en nesten fullstendig lagrekke fra devon til tertiær. Flere ganger i dette tidsrommet var området tørt land med vegetasjon og sumpskoger som dannet kullag. I lag fra tidlig og midtre devon på Nord-Spitsbergen er det mange godt bevarte fossiler av tidlige landplanter. Studier av dem har betydd mye for forståelsen av jordas planteliv tidlig i devonperioden. I lag fra sen devon på Bjørnøya finnes en flora av store, treformete karsporeplanter (Lepidodendron og andre) og kullag som det var gruvedrift på i perioden 1916-1926. Spitsbergen har avsetninger fra tidlig karbon med en lignende flora og kullag som russerne har utnyttet ved Pyramiden. I slutten av devon til tidlig karbon lå Svalbard på omkring 20-30ºN og hadde et varmt og fuktig klima.
I mesozoikum var avsetningsforholdene i Svalbardområdet mest marine, men det finnes floraer og kullag fra perioder på overgangen mellom trias og jura og i undre kritt. Floraene omfatter bregner, konglepalmer, bennettitter, ginkgoer og bartrær.
Fra den tidlige delen av tertiærperioden kjenner vi rike floraer fra mange lokaliteter over et stort område på Spitsbergen. Fossilene viser at det var frodige blandingsskoger av bartrær og løvtrær. Mange arter tilhører familier som vi i dag kjenner fra vår egen flora og andre floraer i den nordlige tempererte sone. Det var vidstrakte sumpområder hvor skogene ga opphav til mange tykke kullag som det lenge har vært gruvedrift på. På denne tiden lå Spitsbergen på omtrent 70ºN med «mørketid» på vinteren.

Over:
Til venstre: Nålevende Metasequoia fra Botanisk hage, Oslo.
Til høyre: Tertiære fossile planter fra Spitsbergen, bl.a. slektninger av Metasequoia.
Til høyre:
Utsnitt av Sigillaria-stamme fra karbon på Spitsbergen.
|
 |
Systematisk oversikt
I - Organismer som mangler ekte cellekjerne (prokaryoter), éncellete eller trådformete
-
Bakterier; noen former har kjemosyntese; prekambrium - nålevende
-
Blågrønne bakterier, tidligere kalt blågrønnalger; har fotosyntese; prekambrium - nålevende
II - Organismer med ekte cellekjerne (eukaryoter), omfatter de fleste éncellete og alle flercellete organismer
inkludert dyr. (Med unntak av soppene omfatter denne oversikten bare autotrofe eukaryoter med fotosyntese.)
ALGER
Éncellete alger; gruppene som er nevnt her er planktonalger og opptrer som mikrofossiler i marine sedimenter; de enkelte gruppene antas å være eldre enn de eldste fossilene som er kjent til nå.
-
Kalkflagellater (eller coccolitter); karbon - nålevende
-
Fureflagellater (eller dinoflagellater); silur - nålevende
-
Kiselalger (eller diatoméer); tidlig kritt - nålevende
-
Kiselflagellater (eller silikoflagellater); tidlig kritt - nålevende
Makroalger
-
Grønnalger; prekambrium - nålevende (ca. 20 000 arter)
-
Rødalger; kambrium - nålevende (ca. 4 000 arter)
-
Brunalger; devon? - nålevende (ca. 2 000 arter)
SOPP; éncellete og flercellete organismer som er heterotrofe, i symbiose med eller parasitter på andre organismer, eller nedbrytere av dødt organisk materiale; tidlig devon - nålevende (ca. 100 000 arter).
Moser (Bryophyta); silur - nålevende (ca. 25 000 arter)
Karsporeplanter (Pteridophyta); sporebærende planter med ledningsvev.
-
Tre av de tidligste gruppene døde ut i devon: Rhyniofytter (midtre silur - tidlig devon), zosterofyllofytter (tidlig - sen devon), og trimerofytter (tidlig - midtre devon).
-
Progymnospermene, som regnes som forløpere for de nakenfrøete plantene, døde også ut i devon.
De tre nålevende gruppene fantes allerede i devon:
Lepidodendron; trær, sen devon - midtre perm
Sigillaria; trær, tidlig karbon - midtre perm
Calamites; trær, sen devon - tidlig perm
Frøplanter
Nakenfrøete (gymnospermer)
-
Konglepalmer (cycadéer); tidlig karbon - nålevende (ca. 100 arter)
-
Bennettitter; sen trias - sen kritt
-
Frøbregner; sen devon - jura
-
Ginkgoer; sen karbon - nålevende (én art, Ginkgo biloba)
-
Cordaiter; karbon - perm
-
Bartrær; sen karbon - nålevende (ca. 600 arter)
Dekkfrøete (angiospermer); tidlig kritt - nålevende (ca. 250 000 arter)
Ill.: Hans Arne Nakrem & Svein B. Manum