Før vi tar for oss utviklingen av virveldyrenes tenner kan det være nyttig å beskrive en tann hos mennesket. Vi tilhører jo pattedyrene, det nyeste, yngste trinn i utviklingen av virveldyrene. Tennene hos alle pattedyr er likt bygget bortsett fra forskjeller i form.
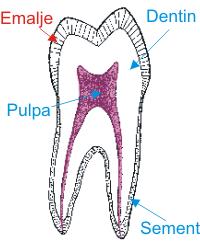 Fig. 1. Snitt gjennom menneskelig tann. Den består hovedsakelig av dentin (tannbein), et hardt, levende vev som får sin næring fra blodårer i pulpa (tannmargen). Der er ingen celler i dentinet slik som i bein, men det inneholder millioner av tynne kanaler med celleutløpere fra pulpa. Utenpå kronedentinet ligger emaljen som er kroppens hardeste vev og skal tåle all slags slitasje ved tygging. Den inneholder hverken celler eller celleutløpere. Utenpå roten ligger sementen som inneholder celler slik som bein i skjelettet. I sementen festes millioner av fibre som i sin andre ende festes i kjevebeinet. Det faste dentin og sementen består av bindevevsfibre og apatitt, dvs. kalsiumfosfat liksom i skjelettet ellers. Tannemalje har ingen fibre men er praktisk talt ren apatitt. Apatitt er karakteristisk for virveldyrs skjelett og tenner, i motsetning til kalsiumkarbonat, kitin og keratinforbindelser som utgjør skjelett og annet hardvev hos insekter, edderkopper, krepsdyr og alle andre virvelløse dyr.
Fig. 1. Snitt gjennom menneskelig tann. Den består hovedsakelig av dentin (tannbein), et hardt, levende vev som får sin næring fra blodårer i pulpa (tannmargen). Der er ingen celler i dentinet slik som i bein, men det inneholder millioner av tynne kanaler med celleutløpere fra pulpa. Utenpå kronedentinet ligger emaljen som er kroppens hardeste vev og skal tåle all slags slitasje ved tygging. Den inneholder hverken celler eller celleutløpere. Utenpå roten ligger sementen som inneholder celler slik som bein i skjelettet. I sementen festes millioner av fibre som i sin andre ende festes i kjevebeinet. Det faste dentin og sementen består av bindevevsfibre og apatitt, dvs. kalsiumfosfat liksom i skjelettet ellers. Tannemalje har ingen fibre men er praktisk talt ren apatitt. Apatitt er karakteristisk for virveldyrs skjelett og tenner, i motsetning til kalsiumkarbonat, kitin og keratinforbindelser som utgjør skjelett og annet hardvev hos insekter, edderkopper, krepsdyr og alle andre virvelløse dyr.
Som alle virveldyr har vi et indre skjelett bygget på ryggraden. Når begynte utviklingen av denne dyregruppen? Den skjedde i havet, lenge før landdyrene oppsto. De første spor av virveldyr fra 500 millioner år siden, er pussig nok tenner, eller rettere sagt et gripeapparat av apatitt med 0,25 til 2 mm. store tannelementer. Disse tannelementene besto av dentin og emalje av apatitt. Først i 1993 ble man klar over at de stammet fra fisk uten kjever eller skjelett for øvrig, selv om dyrene anatomisk må defineres som virveldyr; bl. a. fant man at de hadde ryggmarg, det vil si den store hovednerve på ryggsiden.
Etter hvert utviklet de tidligste virveldyrene et ytre skjelett. Det var de kjeveløse panserfisk. Disse dominerte virveldyrene gjennom et tidsrom på 150 mill. år, fra 500 til 350 mill. år siden. Panseret deres besto av et ytre skjelett av beinplater, placoidskjell, som spesielt dekket hode og forkropp i et tett mønster, og sannsynligvis også avstivet munnåpningen og kunne tjene som griperedskap mot byttedyr. Mikroskopisk og kjemisk er beinet i placoidskjell likt dentinet i våre tenner. Det er mer kompakt og hardere enn knokkelvev i skjelettet. Placoidskjell var ikke dekket av emalje men der var pulpahulrom eller kanaler med blodkar og celler i dette dentinet.
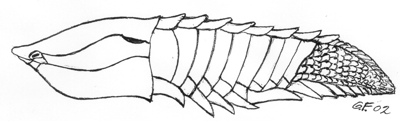 Fig. 2. Gjennom et tidsrom på 150 millioner år gjennomgikk de kjeveløse panserfisk økende utbredelse og artsrikdom.. De var alle tannløse representanter for de tidligste virveldyr oghadde et ytre skjelett av større eller mindre tettsittende plater av dentin, placoidskjell. Tegningen viser en Anglaspis fra tidlig devon; ca 412 mill. år siden. Dette eksemplaret var bare 11 cm langt.
Fig. 2. Gjennom et tidsrom på 150 millioner år gjennomgikk de kjeveløse panserfisk økende utbredelse og artsrikdom.. De var alle tannløse representanter for de tidligste virveldyr oghadde et ytre skjelett av større eller mindre tettsittende plater av dentin, placoidskjell. Tegningen viser en Anglaspis fra tidlig devon; ca 412 mill. år siden. Dette eksemplaret var bare 11 cm langt.
Mot slutten av den nevnte perioden oppsto etter hvert kjever fra de fremste gjellebuer samtidig som det ytre panserskjelett fikk mindre betydning hos virveldyr. Hos disse kjevemunnede virveldyr som hadde et indre skjelett av brusk, utviklet placoidskjell på kjevene seg til tenner. Det skjedde antakelig på samme måte som vi ser det hos hai i dag.
Hos moderne haiarter som ellers har bevart mange andre primitive trekk fra sine opprinnelige former i sen devon (350 mill. år siden), er likhetene mellom tennene og de små tettliggende placoidskjell på kroppens overflate tydelige i mikroskopet, både i form og ved sin utvikling og vekst. Placoidskjell oppstår akkurat som tenner, som et produkt mellom bindevevet og hud eller slimhinne, dvs. epitelet over, som fig. 3 viser for en haitann. På visse steder mellom de to vevstyper former grenselinjen seg til omrisset av den tann (eller placoidskjell) som skal dannes der og høye bindevevsceller like under epitelet avsetter dentin lagvis. Disse høye bindevevsceller kalles odontoblaster. Epitelcellene nærmest dentinet er også høye sylinderceller. De deltar ikke direkte i dannelsen av hardvev hos fisk, frosk, padde og salamander, men langt senere i virveldyrenes utvikling produserer de emalje. Liksom bein i skjelettet, inneholder dentin større hulrom og kanaler fylt av celler og blodkar, se fig. 4. Utenom celler og blodkar består det kompakte dentin av apatitt krystaller i en tett vev av fibre. På spissen og overflaten av tannen er dentinet hardere enn ellers; det kalles vitrodentin (vitro- = glass-) og er også dannet av bindevevscellene.
For å se hvordan et organ er bygget opp, må vi skjære tynne snitt av det, farge disse og se på dem i mikroskop.
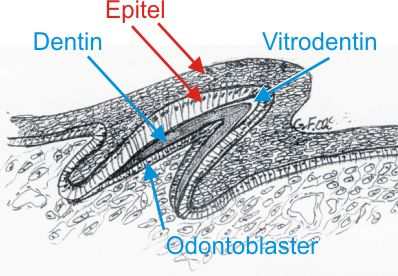 |
Fig. 3 Viser skjematisk mikroskopsnitt gjennom tanndannelse (eller placoidskjelldannelse) hos moderne hai. Avsettelsen av dentin fra odontoblastene er kommet i gang. De høye sylinderceller i epitelet mot dentinet avsetter ikke hardvev hos fisk, salamander eller frosk, men i virveldyrenes utvikling mot pattedyr vil de produsere emalje. Den klare sone på tannspissen er vitrodentin som også er et bindevevsprodukt liksom dentinet ellers. Utviklingsmessig er vitrodentin helt ulikt vår tannemalje. |
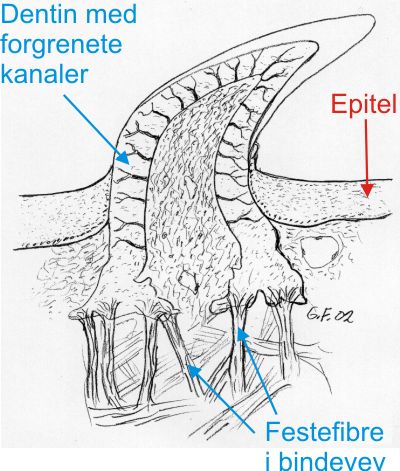 |
Fig 4. Mikroskopsnitt som viser den ferdige, frembrudte tann hos hai med blodkar og celler i de forgrenede kanaler som stråler ut i dentinet, og det hardere celle- og blodkarfrie vitrodentinet på spissen og i et tynt lag på tannoverflaten. Haitenner har ikke røtter festet i kjevebrusken, men derimot sterke og tallrike fibre som feste i bindevevet mellom tann og kjeve. Haitenner har ikke endret seg mye på 350 mill. år. |
Hos primitive virveldyr som hai, er tennene enkle, like og koniske av form, i blant med sagtannede kanter eller, hos noen haiarter, flate for knusing av skjelldyr. Tennene felles og veksles gjennom hele livet, se fig. 5.
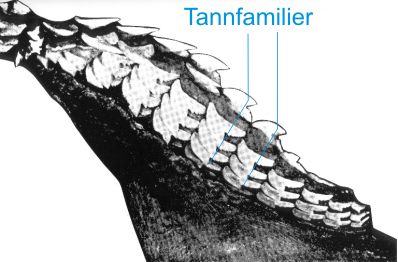 |
Fig. 5. Haikjeve hvor slimhinnen og bindevevet på innsiden mot munnhulen er fjernet så man kan se de nye, ikke frembrudte tenner. Disse er ordnet i "familier" som er tannrekker på tvers av kjeven. Hver familie ligger forskjøvet en halv tannhøyde i forhold til de to nabofamiliene. Til enhver tid er bare den øverste tannen i annen hver familie i funksjon. |
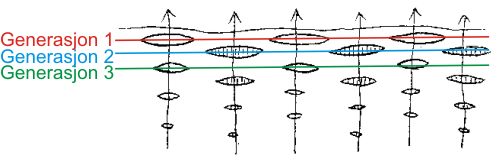 |
Fig. 6. Skjematisk fremstilling av tannsett hos hai.Pilene viser frembruddsretningen for tennene. I annen hver familie utgjør alle tenner av samme alder én tanngenerasjon. Her er den eldste generasjonen angitt med rødt, nest eldste med blått og neste med grønt. De tre yngre generasjonene er ikke markert. |
|

|
Fig.7. Tverrsnitt gjennom underkjeve og tennene i én tannfamilie hos hai. Betingelsen for et kontinuerlig tannskifte er den tynne tannlisten av epitel som strekker seg ned fra fungerende tannrekke på innsiden av kjeven i bindevevet mellom munnepitelet og kjeven. På denne listen oppstår fortløpende nye tannanlegg, og de flytter seg mot de fungerende tenner etter som disse felles. |
Hos moderne frosk, padder og salamandre og noen nålevende samt de fleste fossile reptiler eksisterer et slikt tannvekslingsmønster som vist i figurene 5 og 6. Det er også karakteristisk at alle tennene er like.
Hos beinfisk som torsk, gjedde osv. er tennene også oppbygget av dentin med vitrodentin på tannspissen, og tenner veksles og nydannes hele livet. Imidlertid er det vanlig at hver tann er stivt festet til en beinsokkel som igjen er fast forankret til kjevebeinet. Beinet i denne sokkelen er forskjellig fra kjevebeinet og dentinet og forsvinner etter at tannen er felt. Den yngre erstatningstannen får dannet en ny sokkel ved skiftet, se fig. 8.
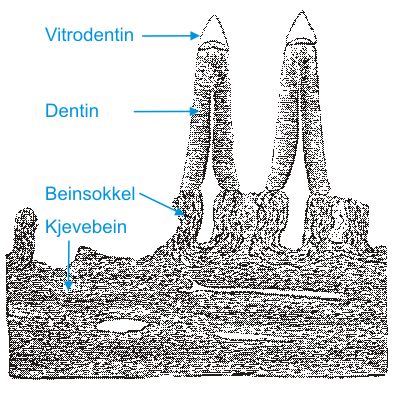 |
Fig.8. Tannsettet hos beinfisk består av like tenner som hos hai. Tannfestet karakteriseres oftest ved fast forbindelse mellom kjevebein og tann via en beinsokkel som nydannes ved tannskiftet. Bein i sokkel og kjeve har forskjellig oppbygning og i tegningen er også grenselinjen mellom de to vev markert. Forbindelsen mellom tann og sokkel brytes lett, så tannskifte er omtrent like hyppig som hos hai. |
Hos torsk er der langs kjeven tallrike uregelmessige rekker av funksjonerende tenner. Tannspissene stikker opp gjennom epitelet og nye, ennå ikke festede anlegg dannes fra dette epitel nær tenner som snart skal felles i hver rekke. Når fellingen skjer er en ny tann klar og dens sokkel og feste til kjevebeinet blir dannet. Tannveksling og -feste hos beinfisk er altså ganske forskjellige fra forholdene hos hai og mange andre bruskfisk. De største tennene i overkjeven sitter ytterst; de minker gradvis gjennom rekkene mot ganen, mens de største i underkjeven alltid er i den innerste tannrekken mot tungen. Tennenes absolutte størrelse står i forhold til fiskens størrelse. Der er dessuten alltid færre rekker i under- enn i overkjeven.
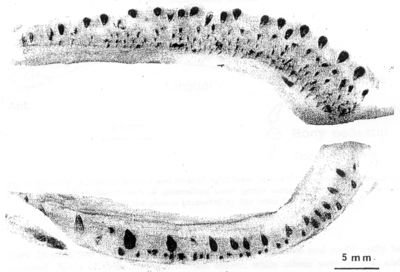 |
Fig. 9. Høyre halvdeler av torskekjever, overkjeve (øverst) og underkjeve, med uregelmessige rekker av frembrutte tenner. Nesten alle de synlige tennene er i funksjon, men noen få nye er ennå ikke festet i kjevebeinet. I overkjeven sitter de største tennene ytterst, i underkjeven innerst. |
Pattedyrenes sannsynlige utviklingshistore
Systemet med mange like tenner direkte festet til kjevebeinet og et livslangt tannskifte dominerte også hos reptiler (krypdyr) fra karbon til slutten av krittiden, dvs. gjennom 300 mill. år. De fleste nålevende reptiler har også slike tannsett. Aligator og krokodille er eneste unntak mht. tannfeste, siden de har en elastisk, sterk forbindelse mellom tann og kjeve liksom pattedyrene, se fig. 16.
Fra perm og til slutten av trias levde de pattedyrlignende reptiler, se fig. 10. Deres kjennetegn var en mer oppreist gange enn hos andre reptiler, utvikling og forenkling av kjeveleddet og effektivisering av tyggemuskulatur og sterk reduksjon i antall tenner i tannsettet. Tennene kan inndeles i karakteristiske grupper med fortenner, hjørnetenner og jeksler. Antall tannskifter ble også redusert og tennenes feste til kjeven lignet de senere pattedyrs. Det er mulig at kroppstemperaturen hos disse reptilene ikke varierte så mye med luftens temperatur som hos tidligere, eller andre samtidige, mer primitive reptiler. Man mener at huden hos enkelte former var hårkledd. Ved overgangen til jura, se fig. 10, forsvant disse avanserte reptiler. Gjennom jura- og krittiden var landdyrene dominert av andre reptilformer, inklusive dinosauriene. Noen av disse var de største landdyr som har eksistert. I dette mangfoldet av reptiler finner vi samtidig de primitive pattedyr som ennå var svært små, dvs. lik eller mindre enn en rotte. Hos alle primitive pattedyr fra tidlig Jura, har vi ett eneste tannskifte som bare omfatter incisiver, hjørnetenner og premolarer. Jekslene ble aldri skiftet. Tannemalje som produseres av de sylindriske epitelcellene mot dentinet, se fig 3, eksisterte alt hos de pattedyrlignende reptiler. Men det er først hos primitive pattedyr, særlig i krittiden, at emaljens mikroskopiske bygning begynner å ta samme form som hos nålevende pattedyr.
Våre moderne pattedyr omfatter tre grupper:
1: monotremater som er merkelige, primitive pattedyr i Australia og på Tasmania, Ny Guinea og New Zealand og består av nebbdyret og to arter insektetende pinnsvin. Begge legger egg og mangler tenner. Monotrematene er truede dyrearter.
2: pungdyr (marsupialia) som omfatter mange slekter og arter i Australia, på Tasmania og New Zealand og noen få i Nord- og Sydamerika (kenguru tilhører den mest kjente slekten i Australia). Etter et kort svangerskap fødes ungene som er svært små og lite utviklet. De må derfor tilbringe lang tid i morens pung under resten av utviklingen.
3: placentalia som vi selv tilhører og som er den dominerende gruppe i antall arter og individer og som fins på alle kontinenter. Fordi gravide mødre har morkake (placenta) er svangerskapet langt og ungene fødes nesten fullt utviklet.
Der er ingen fundamental forskjell mellom pungdyr og vanlige placentalia når det gjelder tennenes form og mikroskopiske bygning. En eiendommelighet hos pungdyr er at bare én tann i hver kjevehalvdel skiftes, og det er 3. premolar. I hele tannsettet er det altså bare 4 melketenner som skal skiftes før dyret blir voksent! En annen eiendommelighet er at det er flere tannanlegg i kjeven enn det er tenner som kommer i funksjon. Disse overskytende tannanleggene forsvinner før tannfrembruddet og man vet ikke om de representerer tidligere tannsett som er tilbakedannet under pungdyrenes utvikling. Deres stilling i kjeven samsvarer imidlertid ikke med anlegget til den ene melketann som finnes for 3. premolar. Hos placentalia skiftes alle tenner foran jekslene og alle tannanlegg utvikles til fungerende tenner.
|
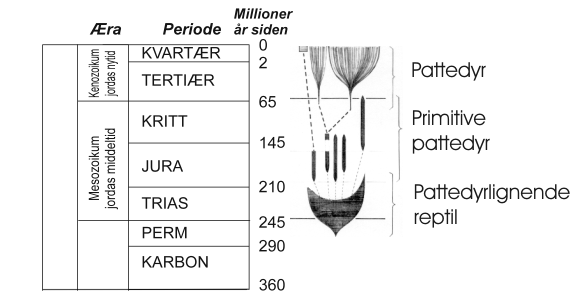
Fig. 10. Fra begynnelsen av perm til i dag representerer skjemaet 290 mill. år. Det er i løpet av dette tidsrom utviklingen mot pattedyrene foregikk. |
Den basale tannformel hos pungdyr og placentalia
Hos fisk, amfibier og reptiler er der et høyt antall like tenner som veksles hyppig. Opprettholdelse av tannsettet som helhet krever mye energi for stadig nydannelse av anlegg og nedbryting og gjenoppbygning av tannfester. Hos de pattedyrlignende reptiler i perm trias (se fig. 10) var som tidligere beskrevet antall tenner redusert betydelig og de var ordnet i forskjellige grupper mht. form og funksjon.. Til sist i utviklingen har vi pattedyrene hvor antall tenner har ett maksimum hos pungdyr og et annet hos placentalia som vist i tannformlene i figurene 11 og 12 nedenfor. De forskjellige tanngrupper har latinske navn og første bokstav i disse angir tanngruppen. Der var fortenner (incisiver)) til avbiting, lange og spisse hjørnetenner (caniner) for avliving av byttedyr, mindre kinntenner med to tyggeknuter (premolarer) og endelig større jeksler med mange tyggeknuter (molarer) for knusing og oppdeling av føden. Bare en del av tannsettet hos pattedyr skiftes. De store jekslene skiftes aldri. Hos pattedyr produserer de høye sylindercellene i epitelet emalje utenpå kronedentinet, se fig. 3. Dette skjedde også hos avanserte reptiler,.
 |
Fig. 11. Den amerikanske opossum har det maksimale antall tenner hos pungdyr. Den hører kanskje til de minst spesialiserte av pungdyrene. Sammenlign med den placentale tannformelen hos hund nedenfor. |
 |
Fig 12. Hund har beholdt alle tenner, unntatt siste jeksel i overkjeven i forhold til den basale, placentale tannformel. |
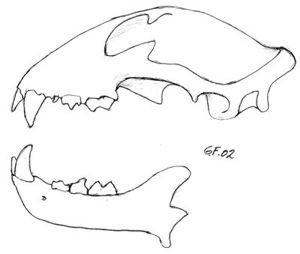 |
Fig. 13. Tannformel hos kattedyr er: 3/3 I 1/1 C 3/2 P 1/1 M og er altså kraftig redusert fra den basale tannformel. |
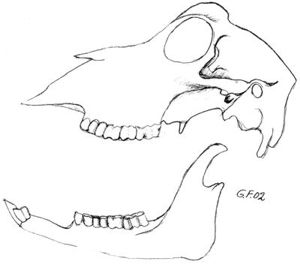 |
Fig. 14. Tannsettet hos geit. Drøvtyggere mangler alle 4 hjørnetenner og fortennene i overkjeven. På tyggeflatene har jekslene skarpe lister av emalje, slik at disse tennene egner seg spesielt til knusing og maling av plantedeler. |
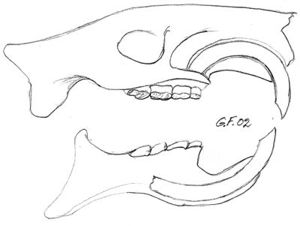 |
Fig. 15. Beverkranium hvor benet over incisivene er fjernet. Hos gnagere slites de lange krumme incisiver like fort som de bryter frem. Tegningen viser at incisivenes rot ligger under jekslenes røtter. Ingen gnagere har hjørnetenner. Noen gnagere har to fortenner i hver kjevehalvdel i stedet for bare én, f. eks. hare. |
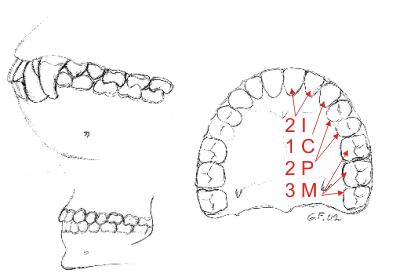 |
Fig. 16. Øverst tannsettet hos gorilla, under hos menneske i profil. Mennesket har samme tannformel som menneskeapene: 2/2 I 1/1 C 2/2 P 3/3 M. Men siste jeksel, visdomstannen, bryter ikke alltid frem hos oss og den vil med tiden forsvinne på samme måte som tenner hos andre pattedyr har forsvunnet. Det er ellers tydelig at hjørnetennene er størst og kraftigst hos menneskeapene. Nederst tannformel for venstre overkjeve hos menneske. Hos menneskeaper og mennesket er ennå tannsettet lite redusert og spesialisert i forhold til den basale tannformel, sammenlign med figurene 12 15.
|
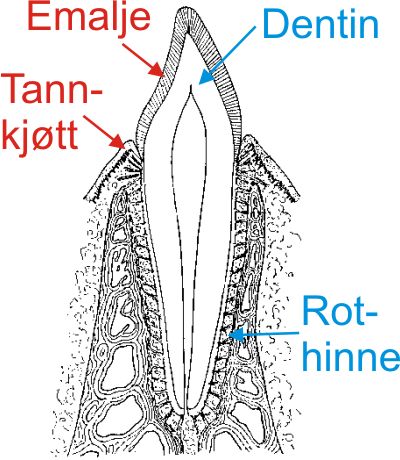 |
Fig. 17. Pattedyrtenner er festet i dype alveoler i kjevene ved hjelp av en kraftig rot og en senehinne mellom rot og bein. Senehinnen som kalles rothinne, består av bindevevsfibre festet i sementen som ligger utenpå rotdentinet, og på den andre siden i beinplaten inne i alveolen. Kronedentinet er dekket av en tykk emaljekappe. Den er dannet av epitelet som opprinnelig trekker opp tannens form, se fig. 3. |
 |
Fig. 18 Tannformene hos levende eller fossile pattedyr er særegne for hver art. Disse tennene er fra pattedyr som levde i krittiden. Spesielt er arrangementet av tyggeknutene helt typisk i de minste detaljer. Dette gjør identifiseringen av fossile arter langt sikrere enn ved hjelp av andre skjelettdeler. Dessuten er emaljen også det hardvev som sist ødelegges. Dette er gunstig for oss fordi emaljens mikroskopiske oppbygning er typisk for visse grupper av pattedyrarter. Men den direkte synlige tannformen er det nøyaktigste kjennemerke for en fossil pattedyrart. Av disse grunner har man bra oversikt over de forskjellige pattedyrformer som oppsto og forsvant og de former som besto og utviklet seg videre mot større artsrikdom inntil moderne tid. Hver tann i montasjen er vist fra siden og ovenfra. Tenner nr. 1-4 er fra arter av en pattedyrorden som døde helt ut for ca 60 mill. år siden. Tenner nr. 5 og 6 er også fra utdøde pattedyrarter, men de var forløpere for tallrike nålevende former. Nr. 5 var forløper for moderne insektetere: f. eks. pinnsvin. Nr. 6 var forløper for bl. a. moderne drøv-tyggere, f. eks. geit. Særlig drøvtyggeres tenner har endret seg mye fra denne fossile formen. |
Som vi har sett er dentinets mikroskopiske struktur og kjemiske oppbygning hos virveldyrene mye eldre enn oppkomsten av de første bindevevsfestede tenner med dentin i sen devon. Tenner bygget på dentin med en kappe av hard emalje på kronen i tillegg, kunne motstå langvarig slitasje uten veksling når de dessuten hadde et godt feste i kjeven. Dette var mer økonomisk enn et stadig tannskifte. Tennene fikk dessuten former som var tilpasset forskjellige typer mat hos spesialiserte planteetere eller kjøttetere. Under utviklingen var også kjeveleddet forenklet og styrket sammen med endringer i tyggemuskulaturen. Skarpe tenner som kan dele opp og finmale maten, effektiv tygging, gir en bedre utnyttelse av matens energiinnhold og er en nødvendighet for utvikling av varmblodighet og et høyt stoffskifte- og energinivå hos pattedyrene. De kunne av disse grunner føre et aktivt liv også i tempererte og kjøligere omgivelser under skiftende årstider . Samtidig ble hjernen større og mer komplisert; intelligens er også en nødvendig forutsetning for å mestre naturens utfordringer. Til sist, hos mennesket, er det hjernens kapasitet som er det største fortrinn i konkurransen om å utnytte og beherske naturen. Men tannutviklingen representerer altså viktige skritt på veien under denne utvikling av virveldyrene.
Faktaside om tenner og tannfunn, vanlige norske pattedyr: http://natmus.uio.no/geologi/faktablader/blad45.htm
Ill.: Gisle Fosse